The genealogical research method is used to establish the dominant
Genealogical method
F. Galton's first work on the genetics of psychological traits is devoted to the analysis of the pedigrees of outstanding people. In other words, it uses the genealogical method, i.e. family research method. It is based on simple logic: if any trait is encoded in genes, then the closer the relationship (i.e., the more identical genes), the more similar these people should be to each other in terms of this trait.
A prerequisite for using the genealogical method is the presence of relatives of the first degree, forming the so-called nuclear (“nuclear”) family; These include “parent-child” and “sibling-sibling” pairs. They, and only they, have on average 50% of their genes in common. Further, with a decrease in the degree of relatedness, the proportion of common genes decreases, and, according to the method, the similarity of people should decrease. The more generations the family tree includes and the wider the circle of relatives, i.e. The more extensive it is, the more reliable the results obtained should be. The interpretation of the results with such comparisons is the same as in the twin method (Fig. 4.3).
Figure 4.3.
When compiling geneagrams, a certain system of symbols is used (Fig. 4.4). When using geneagrams, the following inheritance options can be identified:
Dominant (for example, brachydactyly, polydactyly, achondroplasia);
Recessive (galactosemia, phenylketonuria);
Gender-linked (color blindness, hemophilia, etc.).
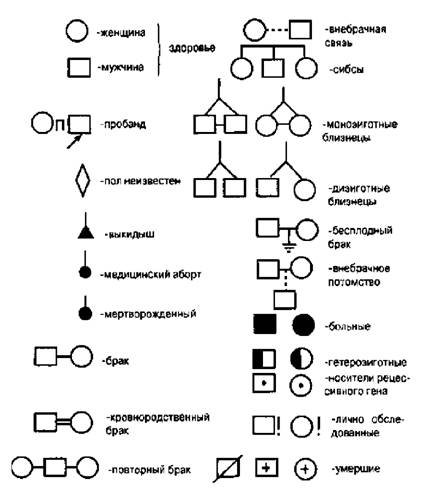
Figure 4.4.
The theoretically calculated kinship coefficients for relatives are also taken into account (Table 4.3).
Table 4.3. Theoretically calculated relatedness coefficients for various pairs of relatives
Very interesting family trees families that gave the world several outstanding scientists and musicians - for example, the Bach family, Bernoulli, etc. In particular, the Bernoulli family is a Swiss Protestant family, many of whose members made significant contributions to science in the 19th-20th centuries. This dynasty includes prominent mathematicians and physicists (3 of them great), as well as famous historians, art critics, architects, lawyers, etc. Historians have counted at least 30 famous representatives of the Bernoulli family in science and culture. Among the academicians of the St. Petersburg Academy of Sciences, five are from the Bernoulli family. The Department of Mathematics at the University of Basel was occupied for 105 years almost without interruption by representatives of the Bernoulli family (Fig. 4.5).
Three generations of Bernoulli produced 9 major mathematicians and physicists, of whom the most famous are:
Bernoulli, Jacob (1654-1708);
Bernoulli, Johann (1667-1748), younger brother Jacob;
Bernoulli, Daniel (1700-1782), son of Johann;
Bernoulli, Jacob II (1759-1789), nephew of Daniel.
Compilation of a pedigree begins with the individual who was examined first - he is called proband. Usually this is an individual who has manifestations of the trait being studied, but this is not necessary. The pedigree is accompanied by a brief entry about each member of the pedigree and an indication of his relationship to the proband. Based on data on the manifestation of the studied mental property in members of the pedigree, using special methods of genetic and mathematical analysis, the problem of establishing the hereditary nature of the trait is solved. If it is established that the trait being studied is of a genetic nature, then at the next stage the problem of establishing the type of inheritance is solved.
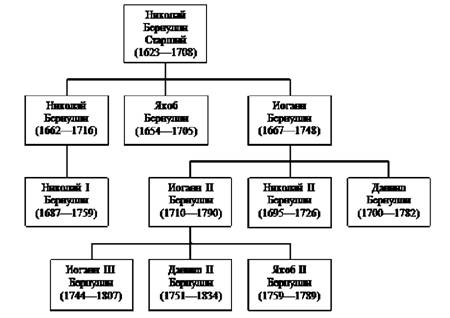
Figure 4.5. Pedigree of mathematicians Return
The clinical and genealogical method is most often used in the genetics of mental illness. Its essence is to trace the manifestations of pathological signs in pedigrees using clinical examination techniques, indicating the type of family ties between family members.
This method is used to establish the type of inheritance of a disease or an individual trait, determine the location of genes on chromosomes, and assess the risk of mental pathology during medical genetic counseling. In the genealogical method, two stages can be distinguished - the stage of compiling pedigrees and the stage of using genealogical data for genetic analysis.
Compilation of a pedigree begins with the person who was examined first, he is called proband. This is usually a patient or an individual who has manifestations of the symptom being studied (but this is not necessary). The pedigree must contain brief information about each family member indicating his relationship to the proband. The pedigree is presented graphically using standard notation, as shown in Fig. 16. Generations are indicated in Roman numerals from top to bottom and placed to the left of the pedigree. Arabic numerals denote individuals of the same generation sequentially from left to right, with brothers and sisters, or sibs, as they are called in genetics, arranged in order of their date of birth. All members of the pedigree of one generation are arranged strictly in one row and have their own code (for example, III-2).
Based on data on the manifestation of a disease or some property being studied in members of a pedigree, using special methods of genetic and mathematical analysis, the problem of establishing the hereditary nature of the disease is solved. If it is established that the pathology being studied is of a genetic nature, then at the next stage the problem of establishing the type of inheritance is solved. It should be noted that the type of inheritance is established not by one, but by a group of pedigrees. A detailed description of the pedigree is important for assessing the risk of pathology in a specific member of a particular family, i.e. when conducting medical genetic counseling.
When studying differences between individuals on any trait, the question arises about the causal factors of such differences. Therefore, in the genetics of mental illnesses, the method of assessing the relative contribution of genetic and environmental factors to interindividual differences in susceptibility to a particular disease is widely used. This method is based on the assumption that the phenotypic (observed) value of a trait in each individual is the result of the influence of the individual’s genotype and the environmental conditions in which its development occurs. However, it is almost impossible to determine this in a specific person. Therefore, appropriate generalized indicators are introduced for all people, which then make it possible, on average, to determine the ratio of genetic and environmental influences on an individual. This problem is solved by introducing such a statistical indicator as the dispersion of a trait, which in genetics is called phenotypic variance(Vp).
Rice. 16. Graphic representation of the pedigree, accepted in genetics.
A - example of a family of 3 generations (explanation in the text); B - basic designations used in pedigrees.
Phenotypic variance can be presented as the sum of two variances, one of which characterizes the diversity due to the influence of genetic factors (V G), and the other - the influence of environmental factors (V E):
V P = V G + V E .
The above indicators are associated with such concepts as the indicator of “genetic diversity” (G):
and the indicator of “environmental diversity” (E):
In these formulas, the symbols represent the first letters English words: V - variation, P - phenotypic, G - genetic, E - environmental.
In many cases, not only the overall assessment of the role of genetic and environmental factors, but also the individual components of variance due to such factors are of significant interest.
The genetic component is usually divided into a component that characterizes the influence of individual gene alleles (G A) - an additive genetic component, or heritability coefficient (h 2), and the influence of pairs of alleles, which characterizes intralocus interaction - a dominant genetic component (G D ).
The environmental component of the dispersion can also be presented in the form of several components, or components. First of all, the variance component is isolated, which is due to the influence of the environment and acts in a similar way on a group of individuals. This is the influence of the so-called systematic (common) environmental factors (E c), which in turn can also be divided into separate types. Another group of effects of environmental factors is characterized by the fact that they act on an individual randomly; the corresponding component of the environmental dispersion is denoted E w .
The considered indicators of genetic and environmental determination, constituting the most important part of genetic analysis in psychiatry, however, do not fully answer the question of the influence of genetic and environmental factors on the manifestation of a trait in a particular person. Thus, if it is established that the diversity of a trait is determined primarily by genetic factors, then this indicates the existence of genetic mechanisms for determining the disease or trait, and the opposite statement is not always true. For example, if a group of subjects is represented by individuals with the same genotype, then there will be no genotypic diversity and, accordingly, the heritability indicator will be equal to 0. Thus, the indicator of genetic determination reflects the influence of genetic factors on interindividual diversity, and not in general on the presence of genetic mechanisms of determination of traits . Heritability rates characterize a population, and for the same disease or trait they can have different values depending on specific differences in population structure, for example depending on differences in genotype frequencies.
In the genetics of mental illness, the main approach for assessing the influence of genetic and environmental factors on interindividual differences is the analysis of correlations between relatives. Methods for calculating coefficients [Lilin E.T., Trubnikov V.I., Vanyukov M.M., 1984] depend on the nature of the distribution of phenotypic values of the analyzed traits (quantitative, alternative nominal-dichotomous or quasi-continuous) and the type of relatives.
Based on the concept of gene identity by descent, the relationship between the genetic components of variance in correlations between relatives has been established. For correlation in parent-child pairs (R OP) (abbreviation “or” from the English offspring (descendant, child) and parent (parent); “sb” - from siblings (brothers and sisters)) there is the following relationship: R OP =
The studies yielded results on susceptibility to a number of mental illnesses. The correlation coefficient between first-degree relatives for susceptibility to schizophrenia, according to the latest data, is 0.35, and accordingly, the heritability of susceptibility to schizophrenia is quite high - 70%. The results of studies on the genetic determination of susceptibility to epilepsy using different criteria for determining the epilepsy phenotype also indicate the high heritability of this disease (50-78%) [Ritsner M.S., 1985]. A high heritability coefficient was also obtained for susceptibility to affective psychoses (70%). At the same time, the contribution of genetic factors to susceptibility to manic-depressive psychosis is 76%, and to depression - 46%.
In general genetics, facts have accumulated indicating the influence of gender on the inheritance of a particular disease or trait. We are talking about the unequal participation of female and male gametes in the formation of the zygote and the body as a whole. These differences are explained by the unequal amount of cytoplasm in the egg and sperm; in the latter there is less of it, which in a number of ways determines the greater influence of the mother, i.e. greater similarity of offspring to the mother (matrolineage) than to the father. It is believed that this influence is based on the following reasons: 1) transmission through the cytoplasm of various cell symbionts (often viruses), capable of reduplicating and, as a result, imitating cytoplasmic inheritance; 2) randomness and uneven distribution of cytoplasmic elements associated with hereditary structures (mitochondria, centrioles) among daughter cells; 3) features of the cytoplasm itself, which can arise both under the influence of the external environment (environmental predetermination) and under the influence of the mother’s genotype (genotypic predetermination).
Associated with the phenomena considered is the widespread concept "maternal effect" Genotypic predetermination is essentially the classical definition of the maternal effect in the “narrow” sense of the words. A characteristic feature of this effect is that it is caused by the action of the mother’s nuclear genes, which change the cytoplasm of the egg before fertilization. As a result, the offspring develops in accordance with the mother's genotype, regardless of its own genotype. This is probably explained by the fact that a significant amount of mRNA accumulates in the egg, which is used in the development of the zygote. In cases of mental illness, the situation is complicated by the fact that the similarity of children with their mother may also be due to the intrauterine and postnatal environment. Therefore, all the causes of matrolineage can be combined into the concept of “maternal effect” in the “broad” sense of the words. For example, there is evidence of maternal influence on the age of onset of Huntington's disease. According to these data, late onset of the disease occurs 2 times more often when the mother is ill than when the father is ill. It has also been shown that children of mothers with epilepsy are 1.5-2 times more likely to develop epilepsy than children of fathers with epilepsy. The difficulty of assessing the presence of a maternal effect is also due to the fact that the difference between relatives may also be due to the possible effect of genes located on the X chromosome, i.e. taking into account their gender.
The specific mechanisms of the maternal effect may be different for each case and require special research. One of these mechanisms may be the mother’s heterozygosity for an allele, the influence of which affects the manifestation of the child’s trait in the form of phenocopy. An example of such a mechanism is brain damage in fetuses of women heterozygous for the phenylketonuria (PKU) gene. Research results indicate that approximately 25% of siblings of probands with PKU have cerebral pathology, which is explained by intrauterine hyperphenylalanemia and is caused by the heterozygous mother effect. In this case, knowledge of the mechanism of the maternal effect allows us to implement special preventive measures to prevent damage to children of heterozygous mothers.
One of the variants of the clinical-genealogical method is study of adopted children. This method is based on the fact that children raised in families of adoptive parents have the genes of their biological parents, and the influence of the environment is determined by the conditions in which the adoptive parents live. Therefore, it is possible to determine the contribution of not only genetic factors to susceptibility to mental illness, but also the contribution of environmental factors (including living conditions, upbringing in families, etc.). Thus, studies carried out taking into account modern methodological requirements on a large sample of adopted children whose mothers had schizophrenia, and a control group of mothers, indicate the predominant contribution of genetic factors to the development of the disease. The heritability coefficient calculated from these data was 70%.
The influence of the family environment on susceptibility to the manifestation of the disease in accordance with the method under consideration can also be determined if the following groups are compared: children of sick parents raised by mentally healthy adoptive parents; children of mentally healthy parents raised by mentally ill adoptive parents; children whose natural and adoptive parents are mentally healthy. Thus, a double correlation in susceptibility to the manifestation of the disease in children whose biological parents are sick and whose adoptive parents are healthy will give the magnitude of the contribution of genetic factors to susceptibility to the disease. The correlation in susceptibility to the manifestation of the disease for children whose biological parents are healthy and whose adoptive parents are sick indicates the influence of the postnatal family environment on susceptibility to the disease. The influence of the intrauterine environment on susceptibility to disease can be obtained using parent-child pairs in families where the adoptive parents are healthy and the biological parents are sick.
Considering the possible role of genes located on the X chromosome, the estimate of the influence of intrauterine factors is equal to the difference between mother-son and father-daughter correlations. An assessment of the maternal effect on susceptibility to the manifestation of mental pathology can also be obtained by studying half-siblings, i.e. children, one of whose parents is common and the other is not. There are half-sibs who have the same mother and different fathers, and half-sibs who have the same father and different mothers. Comparison of groups of half-siblings with a common mother or father, with the presence or absence of mental pathology in the parents, makes it possible to establish the influence of intrauterine factors on the occurrence of mental pathology in children. Taking into account the possible effect of the X chromosome, such an estimate can be obtained as the difference between the correlations of male half-sibs on the maternal side and half-sibs of males on the paternal side, when the mother and father, respectively, are sick.
These techniques are simple enough to evaluate the prospects of further specially organized studies to study the role of the maternal effect on the phenotypic manifestation of the trait.
Genealogical method
This method is based on the compilation and analysis of pedigrees. This method has been widely used from ancient times to the present day in horse breeding, selection of valuable lines of cattle and pigs, in obtaining purebred dogs, as well as in breeding new breeds of fur-bearing animals. Human genealogies have been compiled over many centuries regarding the reigning families of Europe and Asia.
As a method of studying human genetics, the genealogical method began to be used only from the beginning of the 20th century, when it became clear that the analysis of pedigrees, which trace the transmission from generation to generation of a certain trait (disease), can replace the hybridological method, which is actually inapplicable to humans.
When compiling pedigrees, the starting point is the person - the proband, whose pedigree is being studied. Usually this is either a patient or a carrier of a certain trait, the inheritance of which needs to be studied. When compiling pedigree tables, the symbols proposed by G. Just in 1931 are used (Fig. 6.24). Generations are designated by Roman numerals, individuals in a given generation are designated by Arabic numerals.
Rice. 6.24. Legend when compiling genealogies (according to G. Just)
Using the genealogical method, the hereditary nature of the trait under study can be established, as well as the type of its inheritance (autosomal dominant, autosomal recessive, X-linked dominant or recessive, Y-linked). When analyzing pedigrees for several characteristics, the linked nature of their inheritance can be revealed, which is used in the compilation of chromosomal maps. This method allows you to study the intensity of the mutation process, assess the expressivity and penetrance of the allele. It is widely used in medical genetic counseling to predict offspring. However, it should be noted that genealogical analysis becomes significantly more complicated when families have few children.
Pedigrees with autosomal dominant inheritance. The autosomal type of inheritance is generally characterized by an equal probability of occurrence of this trait in both men and women. This is due to the same double dose of genes located in the autosomes of all representatives of the species and received from both parents, and the dependence of the developing trait on the nature of the interaction of allelic genes.
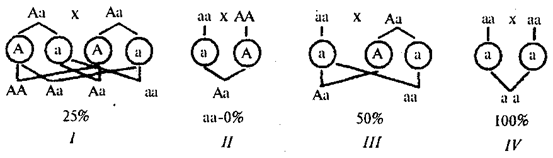
When a trait dominates in the offspring of a parental pair, where at least one parent is its carrier, it appears with greater or lesser probability depending on the genetic constitution of the parents (Fig. 6.25).
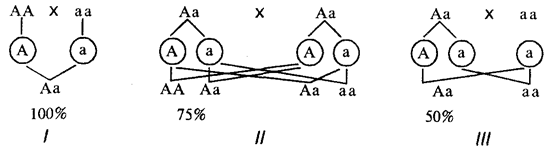
Rice. 6.25. Probability of offspring with a dominant trait from different married couples (/- III)
If a trait is analyzed that does not affect the viability of the organism, then carriers of the dominant trait can be both homo- and heterozygotes. In the case of dominant inheritance of some pathological trait (disease), homozygotes, as a rule, are not viable, and carriers of this trait are heterozygotes.
Thus, with autosomal dominant inheritance, the trait can occur equally in men and women and can be traced when there is a sufficient number of offspring in each vertical generation. When analyzing pedigrees, it is necessary to remember the possibility of incomplete penetration of the dominant allele due to the interaction of genes or environmental factors. The penetrance rate can be calculated as the ratio of the actual number of carriers of a trait to the number of expected carriers of that trait in a given family. It is also necessary to remember that some diseases do not appear immediately from the moment the child is born. Many diseases inherited according to a dominant type develop only at a certain age. Thus, Huntington's chorea clinically manifests itself by the age of 35-40, and polycystic kidney disease also manifests itself late. Therefore, when predicting such diseases, brothers and sisters who have not reached a critical age are not taken into account.
The first description of a pedigree with an autosomal dominant type of inheritance of an anomaly in humans was given in 1905. It traces transmission over a number of generations brachydactyly(short-fingered). In Fig. Figure 6.26 shows a pedigree with this anomaly. In Fig. Figure 6.27 shows a pedigree with retinoblastoma in a case of incomplete penetrance.
Pedigrees with autosomal recessive inheritance. Recessive traits appear phenotypically only in homozygotes for recessive alleles. These traits are usually found in the offspring of phenotypically normal parents who are carriers of recessive alleles. The probability of the appearance of recessive offspring in this case is 25%. If one of the parents has a recessive trait, then the likelihood of its manifestation in the offspring will depend on the genotype of the other parent. In recessive parents, all offspring will inherit the corresponding recessive trait (Fig. 6.28).

Rice. 6.26. Pedigree ( A) with an autosomal dominant type of inheritance (brachydactyly - B)
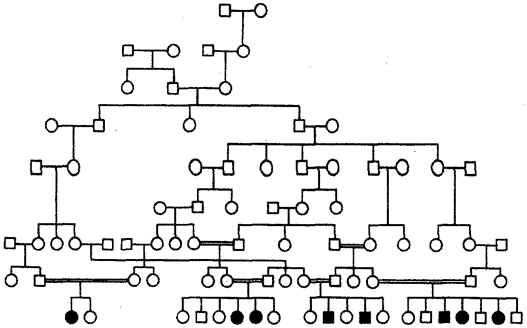
It is typical for pedigrees with an autosomal recessive type of inheritance that the trait does not appear in every generation. Most often, recessive offspring appear in parents with a dominant trait, and the likelihood of such offspring increases in closely related marriages, where both parents may be carriers of the same recessive allele received from a common ancestor. An example of autosomal recessive inheritance is the pedigree of a family with pseudohypertrophic progressive myopathy, in which consanguineous marriages are common (Fig. 6.29). The horizontal spread of the disease in the last generation is noteworthy.
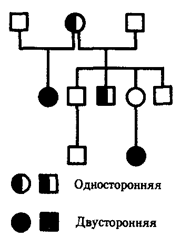
Rice. 6.27. Pedigree with retinoblastoma in case of incomplete penetrance
Rice. 6.28. The probability of producing offspring with a recessive trait
from various married couples ( I-IV)
Pedigrees with dominant X-linked inheritance of the trait. Genes located on the X chromosome and not having alleles on the Y chromosome are present in the genotypes of men and women in different doses. A woman receives her two X chromosomes and corresponding genes from both her father and mother, while a man inherits his only X chromosome only from his mother. The development of the corresponding trait in men is determined by the only allele present in his genotype, while in women it is the result of the interaction of two allelic genes. In this regard, traits inherited in an X-linked manner occur in a population with different probabilities in males and females.
With dominant X-linked inheritance, the trait is more common in women due to the greater possibility of them receiving the corresponding allele either from the father or from the mother. Men can only inherit this trait from their mother. Women with a dominant trait pass it on equally to daughters and sons, while men pass it on only to daughters. Sons never inherit a dominant X-linked trait from their fathers.
Rice. 6.29. Pedigree with an autosomal recessive type of inheritance (pseudohypertrophic progressive myopathy)
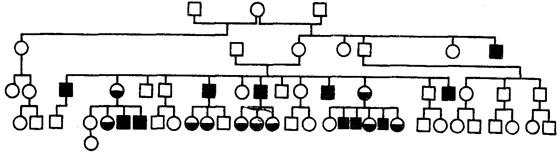
An example of this type of inheritance is the pedigree described in 1925 with follicular keratosis - skin disease accompanied by loss of eyelashes, eyebrows, and scalp hair (Fig. 6.30). The disease is more severe in hemizygous men than in women, who are most often heterozygous.
In some diseases, the death of hemizygous males is observed. early stages ontogeny. Then in the pedigrees among the affected there should be only women, in whose offspring the ratio of affected daughters, healthy daughters and healthy sons is equal to 1: 1: 1. Male dominant hemizygotes that do not die at very early stages of development are found in spontaneous abortions or among stillbirths. These features of inheritance in humans are characterized by pigmentary dermatosis.
Pedigrees for recessive X-linked inheritance of traits. A characteristic feature of pedigrees with this type of inheritance is the predominant manifestation of the trait in hemizygous men, who inherit it from mothers with a dominant phenotype who are carriers of a recessive allele. As a rule, the trait is inherited by men through generations from maternal grandfather to grandson. In women, it manifests itself only in a homozygous state, the likelihood of which increases with closely related marriages.
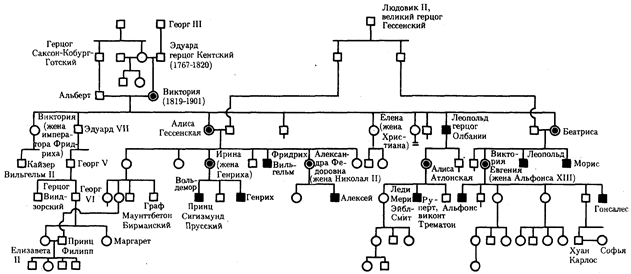
The best known example of recessive X-linked inheritance is hemophilia. The inheritance of hemophilia type A is presented in the pedigree of the descendants of Queen Victoria of England (Fig. 6.31).
Rice. 6.30. Pedigree with X-linked dominant inheritance (keratosis follicularis)
Rice. 6.31. Pedigree for X-linked recessive inheritance (hemophilia type A)
Another example of inheritance by this type is color blindness - a certain form of color vision impairment.
Pedigrees with Y-linked inheritance. The presence of the Y chromosome only in males explains the characteristics of the Y-linked, or holandric, inheritance of the trait, which is found only in men and is transmitted through the male line from generation to generation from father to son.
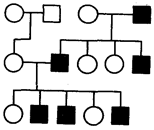
Rice. 6.32. Pedigree with Y-linked (holandric) type of inheritance
One trait whose Y-linked inheritance in humans is still debated is hypertrichosis of the auricle, or the presence of hair on the outer edge of the ear. It is believed that in addition to this gene, the short arm of the Y chromosome contains genes that determine male sex. In 1955, a Y chromosome-determined transplantation antigen called HY was described in the mouse. Perhaps it is one of the factors of sexual differentiation of male gonads, the cells of which have receptors that bind this antigen. The antigen associated with the receptor activates the development of the gonad according to the male type (see section 3.6.5.2; 6.1.2). This antigen has remained almost unchanged in the process of evolution and is found in the body of many animal species, including humans. Thus, the inheritance of the ability to develop gonads according to the male type is determined by the holandric gene located on the Y chromosome (Fig. 6.32).
Twin method
This method consists of studying the patterns of inheritance of traits in pairs of identical and fraternal twins. It was proposed in 1875 by Galton initially to assess the role of heredity and environment in the development of human mental properties. Currently, this method is widely used in the study of heredity and variability in humans to determine the relative role of heredity and environment in the formation of various characteristics, both normal and pathological. It allows you to identify the hereditary nature of a trait, determine the penetrance of the allele, and evaluate the effectiveness of certain external factors (medicines, training, education) on the body.
The essence of the method is to compare the manifestation of a trait in different groups of twins, taking into account the similarities or differences of their genotypes. Monozygotic twins, developing from one fertilized egg are genetically identical, as they have 100% of the same genes. Therefore, among monozygotic twins there is a high percentage concordant couples, in which both twins develop the trait. Comparison of monozygotic twins raised in different conditions postembryonic period, allows us to identify signs in the formation of which environmental factors play a significant role. According to these signs, between twins there is discordance, those. differences. On the contrary, the preservation of similarities between twins, despite the differences in the conditions of their existence, indicates the hereditary conditioning of the trait.
Comparison of pairwise concordance for this trait in genetically identical monozygotic and dizygotic twins, who have on average about 50% of common genes, makes it possible to more objectively judge the role of the genotype in the formation of the trait. High concordance in pairs of monozygotic twins and significantly lower concordance in pairs of dizygotic twins indicate the importance of hereditary differences in these pairs for determining the trait. The similarity of the concordance rate in mono- and dizygotic twins indicates the insignificant role of genetic differences and the determining role of the environment in the formation of the trait or the development of the disease. Significantly different, but rather low concordance rates in both groups of twins make it possible to judge the hereditary predisposition to the formation of a trait that develops under the influence of environmental factors.
Establishing the relative role of heredity and environment in the development of various pathological conditions allows the doctor to correctly assess the situation and carry out preventive measures if there is a hereditary predisposition to the disease or carry out auxiliary therapy if it is hereditary.
The difficulties of the twin method are associated, firstly, with the relatively low frequency of twin births in the population (1:86-1:88), which complicates the selection of a sufficient number of pairs with this trait; secondly, with the identification of monozygosity of twins, which has great importance to obtain reliable conclusions.
A number of methods are used to identify monozygosity of twins. 1. A polysymptomatic method for comparing twins according to many morphological characteristics (pigmentation of the eyes, hair, skin, hair shape and features of hair on the head and body, shape of the ears, nose, lips, nails, body, finger patterns). 2. Methods based on the immunological identity of twins based on erythrocyte antigens (ABO, MN, Rhesus systems) and serum proteins (γ-globulin). 3. The most reliable criterion for monozygosity is provided by a transplantation test using cross-twin skin grafting.
Despite the labor-intensive nature of the twin method and the possibility of errors in determining the monozygosity of twins, the high objectivity of the conclusions makes it one of the widely used methods of genetic research in humans.


















