Ke stanovení dominanta se používá metoda genealogického výzkumu
Genealogická metoda
První práce F. Galtona o genetice psychologických rysů je věnována rozboru rodokmenů vynikajících lidí. Jinými slovy, používá genealogickou metodu, tzn. metoda rodinného výzkumu. Vychází z jednoduché logiky: je-li nějaká vlastnost zakódována v genech, pak čím bližší je vztah (tedy čím více identických genů), tím podobnější by si tito lidé měli být z hlediska této vlastnosti.
Předpokladem pro použití genealogické metody je přítomnost příbuzných I. stupně, tvořících tzv. nukleární („jadernou“) rodinu; Patří mezi ně dvojice „rodič-dítě“ a „sourozenec-sourozenec“. Oni, a jen oni, mají v průměru 50 % společných genů. Dále se snížením stupně příbuznosti klesá podíl společných genů a podle metody by se měla snižovat podobnost lidí. Čím více generací rodokmen zahrnuje a tím širší okruh příbuzných, tzn. Čím je rozsáhlejší, tím spolehlivější by měly být získané výsledky. Interpretace výsledků při takovém srovnání je stejná jako u metody dvojčete (obr. 4.3).
Obrázek 4.3.
Při sestavování geneagramů se používá určitý systém symbolů (obr. 4.4). Při použití geneagramů lze identifikovat následující možnosti dědičnosti:
Dominantní (například brachydaktylie, polydaktylie, achondroplazie);
Recesivní (galaktosemie, fenylketonurie);
Vázané na pohlaví (barvoslepost, hemofilie atd.).
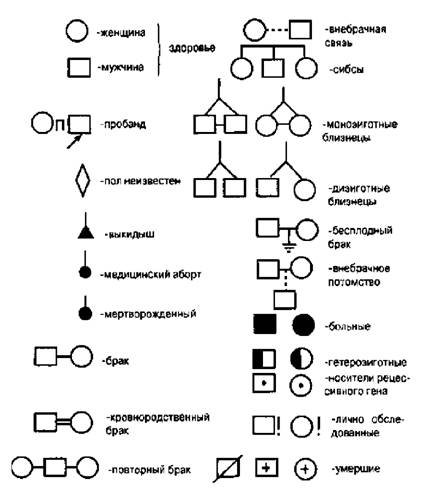
Obrázek 4.4.
Zohledněny jsou i teoreticky vypočtené příbuzenské koeficienty pro příbuzné (tab. 4.3).
Tabulka 4.3. Teoreticky vypočtené koeficienty příbuznosti pro různé dvojice příbuzných
Velmi zajímavé rodokmeny rodiny, které daly světu několik vynikajících vědců a hudebníků – např. Bachovu rodinu, Bernoulliho atd. Zejména rodina Bernoulliových je švýcarská protestantská rodina, jejíž mnozí členové významně přispěli k vědě v 19.–20. století. Do této dynastie patří významní matematici a fyzici (3 z nich velcí), stejně jako slavní historici, kritici umění, architekti, právníci atd. Historici napočítali nejméně 30 slavných představitelů rodu Bernoulli ve vědě a kultuře. Mezi akademiky Petrohradské akademie věd je pět z rodu Bernoulli. Katedru matematiky na univerzitě v Basileji obsazovali téměř bez přerušení 105 let zástupci rodiny Bernoulli (obr. 4.5).
Tři generace Bernoulliho vytvořily 9 hlavních matematiků a fyziků, z nichž nejznámější jsou:
Bernoulli, Jacob (1654-1708);
Bernoulli, Johann (1667-1748), mladší bratr Jacob;
Bernoulli, Daniel (1700-1782), syn Johannův;
Bernoulli, Jacob II (1759-1789), synovec Daniela.
Sestavení rodokmenu začíná u jedince, který byl vyšetřen jako první – je tzv proband. Obvykle se jedná o jedince, který má projevy zkoumané vlastnosti, ale není to nutné. Rodokmen je doplněn stručným záznamem o každém členu rodokmenu a uvedením jeho vztahu k probandovi. Na základě údajů o projevech studovaných duševních vlastností u členů rodokmenu je pomocí speciálních metod genetické a matematické analýzy řešen problém stanovení dědičné povahy znaku. Pokud se zjistí, že studovaná vlastnost je genetické povahy, pak je v další fázi problém stanovení typu dědičnosti vyřešen.
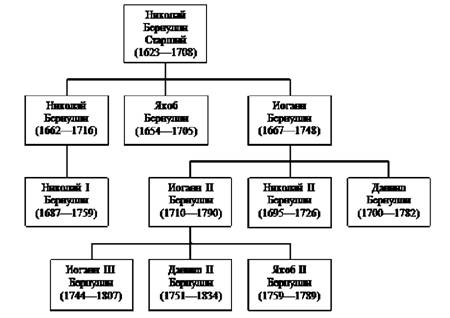
Obrázek 4.5. Rodokmen matematiků Vrátit se
Klinická a genealogická metoda se nejčastěji využívá v genetice duševních chorob. Jeho podstatou je sledování projevů patologických znaků v rodokmenech pomocí klinických vyšetřovacích technik, indikujících typ rodinných vazeb mezi členy rodiny.
Tato metoda se používá ke stanovení typu dědičnosti onemocnění nebo individuálního rysu, určení umístění genů na chromozomech a posouzení rizika duševní patologie během lékařského genetického poradenství. V genealogické metodě lze rozlišit dvě fáze - fázi sestavování rodokmenů a fázi využití genealogických dat pro genetický rozbor.
Sestavování rodokmenu začíná u osoby, která byla vyšetřena jako první, je tzv proband. Obvykle se jedná o pacienta nebo jedince, který má projevy studovaného symptomu (není to však nutné). Rodokmen musí obsahovat stručné údaje o každém členu rodiny naznačující jeho vztah k probandovi. Rodokmen je prezentován graficky pomocí standardní notace, jak je znázorněno na Obr. 16. Generace jsou označeny římskými číslicemi shora dolů a umístěny vlevo od rodokmenu. Arabské číslice označují jednotlivce stejné generace postupně zleva doprava, s bratry a sestrami nebo sourozenci, jak se jim v genetice říká, seřazenými v pořadí podle data narození. Všichni členové rodokmenu jedné generace jsou uspořádáni přísně v jedné řadě a mají svůj vlastní kód (například III-2).
Na základě údajů o projevech onemocnění nebo nějaké vlastnosti studovaných u příslušníků rodokmenu se pomocí speciálních metod genetické a matematické analýzy řeší problém stanovení dědičné povahy onemocnění. Pokud se zjistí, že studovaná patologie je genetické povahy, pak je v další fázi vyřešen problém stanovení typu dědičnosti. Je třeba poznamenat, že typ dědictví není stanoven jedním, ale skupinou rodokmenů. Podrobný popis rodokmenu je důležitý pro posouzení rizika patologie u konkrétního člena konkrétní rodiny, tzn. při provádění lékařského genetického poradenství.
Při studiu rozdílů mezi jednotlivci v jakémkoli znaku vyvstává otázka o kauzálních faktorech takových rozdílů. Proto se v genetice duševních chorob široce používá metoda posuzování relativního podílu genetických a environmentálních faktorů na interindividuálních rozdílech ve vnímavosti ke konkrétnímu onemocnění. Tato metoda je založena na předpokladu, že fenotypová (pozorovaná) hodnota vlastnosti u každého jedince je výsledkem vlivu genotypu jedince a podmínek prostředí, ve kterých k jeho vývoji dochází. Určit to u konkrétního člověka je však téměř nemožné. Pro všechny lidi se proto zavádějí vhodné zobecněné ukazatele, které pak umožňují v průměru určit poměr genetických a environmentálních vlivů na jedince. Tento problém je vyřešen zavedením takového statistického ukazatele, jako je rozptyl znaku, který se v genetice nazývá fenotypový rozptyl(Vp).
Rýže. 16. Grafické znázornění rodokmenu, akceptované v genetice.
A - příklad rodiny 3 generací (vysvětlení v textu); B - základní označení používaná v rodokmenech.
Fenotypový rozptyl lze prezentovat jako součet dvou rozptylů, z nichž jeden charakterizuje rozmanitost způsobenou vlivem genetických faktorů (V G) a druhý - vliv faktorů prostředí (V E):
VP = VG + VE.
Výše uvedené indikátory jsou spojeny s takovými pojmy, jako je indikátor „genetické diverzity“ (G):
a indikátor „environmentální rozmanitosti“ (E):
V těchto vzorcích symboly představují první písmena anglická slova: V - variace, P - fenotypové, G - genetické, E - environmentální.
V mnoha případech je velmi zajímavé nejen celkové posouzení role genetických a environmentálních faktorů, ale také jednotlivé složky rozptylu způsobené těmito faktory.
Genetická složka se obvykle dělí na složku, která charakterizuje vliv jednotlivých genových alel (G A) - aditivní genetická složka, neboli koeficient dědičnosti (h 2), a vliv párů alel, který charakterizuje intralokusovou interakci - dominantní genetická složka. složka (GD).
Environmentální složka disperze může být také přítomna ve formě několika složek nebo složek. V první řadě se izoluje variační složka, která je důsledkem vlivu prostředí a podobně působí i na skupinu jedinců. Jedná se o vliv tzv. systematických (společných) faktorů prostředí (E c), které lze zase rozdělit do samostatných typů. Další skupina působení faktorů prostředí je charakteristická tím, že na jedince působí náhodně; odpovídající složku environmentální disperze označíme Ew.
Uvažované indikátory genetické a environmentální determinace, tvořící nejdůležitější část genetické analýzy v psychiatrii, však plně neodpovídají na otázku vlivu genetických a environmentálních faktorů na projev znaku u konkrétního člověka. Pokud se tedy prokáže, že diverzita znaku je určována především genetickými faktory, pak to svědčí o existenci genetických mechanismů pro určování onemocnění nebo znaku a opačné tvrzení není vždy pravdivé. Pokud je například skupina subjektů reprezentována jedinci se stejným genotypem, pak nebude existovat žádná genotypová diverzita a ukazatel dědičnosti bude tedy roven 0. Ukazatel genetické determinace tedy odráží vliv genetických faktorů na interindividuální diverzitu, a nikoli obecně na přítomnost genetických mechanismů určování znaků . Míry dědičnosti charakterizují populaci a pro stejnou nemoc nebo vlastnost mohou mít různé hodnoty v závislosti na konkrétních rozdílech ve struktuře populace, například v závislosti na rozdílech ve frekvencích genotypů.
V genetice duševních chorob je hlavním přístupem pro hodnocení vlivu genetických a environmentálních faktorů na interindividuální rozdíly analýza korelací mezi příbuznými. Metody výpočtu koeficientů [Lilin E.T., Trubnikov V.I., Vanyukov M.M., 1984] závisí na povaze distribuce fenotypových hodnot analyzovaných znaků (kvantitativní, alternativní nominálně-dichotomické nebo kvazi-spojité) a typu příbuzných.
Na základě konceptu genové identity podle původu byl stanoven vztah mezi genetickými složkami variance v korelacích mezi příbuznými. Pro korelaci v párech rodič-dítě (R OP) (zkratka „nebo“ z anglického potomstvo (potomek, dítě) a rodič (rodič); „sb“ - od sourozenců (bratři a sestry)) platí následující vztah: R OP =
Studie přinesly výsledky o náchylnosti k řadě duševních chorob. Korelační koeficient mezi prvostupňovými příbuznými pro náchylnost ke schizofrenii je podle posledních údajů 0,35, a proto je dědičnost náchylnosti ke schizofrenii poměrně vysoká - 70%. Výsledky studií genetického určení náchylnosti k epilepsii s použitím různých kritérií pro stanovení fenotypu epilepsie rovněž naznačují vysokou dědičnost tohoto onemocnění (50-78 %) [Ritsner M.S., 1985]. Vysoký koeficient dědičnosti byl získán také pro náchylnost k afektivním psychózám (70 %). Současně je příspěvek genetických faktorů k náchylnosti k maniodepresivní psychóze 76% a k depresi - 46%.
V obecné genetice se nashromáždila fakta naznačující vliv pohlaví na dědičnost určité nemoci nebo vlastnosti. Hovoříme o nestejné účasti ženských a mužských gamet na tvorbě zygoty a těla jako celku. Tyto rozdíly jsou vysvětleny nestejným množstvím cytoplazmy ve vajíčku a spermii; v tom druhém je ho méně, což v řadě směrů určuje větší vliv matky, tzn. větší podobnost potomků s matkou (matrolineage) než s otcem. Předpokládá se, že tento vliv je založen na následujících důvodech: 1) přenos cytoplazmou různých buněčných symbiontů (často virů), schopných replikovat a v důsledku toho napodobovat cytoplazmatickou dědičnost; 2) náhodnost a nerovnoměrná distribuce cytoplazmatických prvků spojených s dědičnými strukturami (mitochondrie, centrioly) mezi dceřinými buňkami; 3) rysy samotné cytoplazmy, které mohou vzniknout jak pod vlivem vnějšího prostředí (předurčení prostředí), tak pod vlivem genotypu matky (předurčení genotypu).
S uvažovanými jevy je spojen rozšířený koncept "mateřský efekt" Genotypové předurčení je v podstatě klasickou definicí mateřského efektu v „užším“ slova smyslu. Charakteristickým rysem tohoto účinku je, že je způsoben působením jaderných genů matky, které mění cytoplazmu vajíčka před oplodněním. V důsledku toho se potomstvo vyvíjí v souladu s genotypem matky bez ohledu na jeho vlastní genotyp. To se pravděpodobně vysvětluje tím, že se ve vajíčku hromadí značné množství mRNA, která se využívá při vývoji zygoty. V případech duševního onemocnění je situace komplikovaná tím, že podobnost dětí s matkou může být dána i nitroděložním a postnatálním prostředím. Proto lze všechny příčiny matrolineage spojit do konceptu „mateřského efektu“ v „širším“ smyslu slova. Existují například důkazy o vlivu matky na věk propuknutí Huntingtonovy choroby. Podle těchto údajů se pozdní nástup onemocnění vyskytuje 2x častěji při nemoci matky než při nemoci otce. Bylo také prokázáno, že děti matek s epilepsií mají 1,5–2krát vyšší pravděpodobnost vzniku epilepsie než děti otců s epilepsií. Obtížnost posouzení přítomnosti mateřského efektu je dána i tím, že rozdíl mezi příbuznými může být způsoben i možným vlivem genů umístěných na X chromozomu, tzn. s přihlédnutím k jejich pohlaví.
Specifické mechanismy mateřského efektu se mohou v každém případě lišit a vyžadují speciální výzkum. Jedním z těchto mechanismů může být heterozygotnost matky pro alelu, jejíž vliv ovlivňuje projev dětské vlastnosti ve formě fenokopie. Příkladem takového mechanismu je poškození mozku u plodů žen heterozygotních pro gen fenylketonurie (PKU). Výsledky výzkumu ukazují, že přibližně 25 % sourozenců probandů s PKU má mozkovou patologii, která se vysvětluje intrauterinní hyperfenylanemií a je způsobena efektem heterozygotní matky. Znalost mechanismu mateřského efektu nám v tomto případě umožňuje realizovat speciální preventivní opatření k prevenci poškození dětí heterozygotních matek.
Jednou z variant klinicko-genealogické metody je studium adoptovaných dětí. Tato metoda je založena na tom, že děti vychovávané v rodinách adoptivních rodičů mají geny svých biologických rodičů a vliv prostředí je dán podmínkami, ve kterých adoptivní rodiče žijí. Proto je možné určit podíl nejen genetických faktorů na náchylnosti k duševním chorobám, ale také podíl faktorů prostředí (včetně životních podmínek, výchovy v rodinách atd.). Studie provedené s přihlédnutím k moderním metodickým požadavkům na velkém vzorku adoptovaných dětí, jejichž matky trpěly schizofrenií, a kontrolní skupině matek tedy ukazují na převažující podíl genetických faktorů na rozvoji onemocnění. Koeficient dědičnosti vypočítaný z těchto údajů byl 70 %.
Vliv rodinného prostředí na náchylnost k projevům onemocnění v souladu s uvažovanou metodou lze zjistit i při srovnání následujících skupin: děti nemocných rodičů vychovávané duševně zdravými adoptivními rodiči; děti duševně zdravých rodičů vychovávané duševně nemocnými adoptivními rodiči; děti, jejichž přirození a adoptivní rodiče jsou duševně zdraví. Dvojí korelace ve vnímavosti k manifestaci onemocnění u dětí, jejichž biologičtí rodiče jsou nemocní a jejichž adoptivní rodiče jsou zdraví, tedy dá velikost příspěvku genetických faktorů k náchylnosti k onemocnění. Korelace ve vnímavosti k manifestaci onemocnění u dětí, jejichž biologičtí rodiče jsou zdraví a adoptivní rodiče jsou nemocní, ukazuje na vliv postnatálního rodinného prostředí na vnímavost k onemocnění. Vliv nitroděložního prostředí na náchylnost k onemocnění lze získat pomocí párů rodič-dítě v rodinách, kde jsou adoptivní rodiče zdraví a biologičtí rodiče nemocní.
Vzhledem k možné úloze genů lokalizovaných na X chromozomu je odhad vlivu nitroděložních faktorů roven rozdílu mezi korelacemi matka-syn a otec-dcera. Posouzení mateřského vlivu na náchylnost k projevům duševní patologie lze získat i studiem nevlastních sourozenců, tzn. děti, jejichž jeden z rodičů je společný a druhý ne. Jsou nevlastní sourozenci, kteří mají stejnou matku a různé otce, a nevlastní sourozenci, kteří mají stejného otce a různé matky. Srovnání skupin nevlastních sourozenců se společnou matkou nebo otcem, s přítomností nebo nepřítomností duševní patologie u rodičů, umožňuje zjistit vliv intrauterinních faktorů na výskyt duševní patologie u dětí. S přihlédnutím k možnému vlivu chromozomu X lze takový odhad získat jako rozdíl mezi korelacemi mužských polosourozenců na mateřské straně a polosourozenců mužů na straně otce, kdy matka a otec, resp. , jsou nemocní.
Tyto techniky jsou dostatečně jednoduché na to, aby vyhodnotily vyhlídky na další speciálně organizované studie ke studiu role mateřského účinku na fenotypovou manifestaci znaku.
Genealogická metoda
Tato metoda je založena na sestavování a analýze rodokmenů. Tato metoda se od pradávna až do současnosti hojně využívá při chovu koní, selekci cenných linií skotu a prasat, při získávání čistokrevných psů, ale i při šlechtění nových plemen kožešinových zvířat. Lidské genealogie byly sestavovány po mnoho staletí ohledně vládnoucích rodin v Evropě a Asii.
Jako metoda studia lidské genetiky se genealogická metoda začala používat až od počátku 20. století, kdy se ukázalo, že rozbor rodokmenů, které sledují přenos z generace na generaci určitého znaku (nemoci), může nahradit hybridologickou metodu, která je vlastně pro člověka nepoužitelná.
Při sestavování rodokmenů je výchozím bodem osoba - proband, jehož rodokmen je studován. Obvykle se jedná buď o pacienta, nebo nositele určité vlastnosti, jejíž dědičnost je třeba studovat. Při sestavování rodokmenových tabulek se používají symboly navržené G. Justem v roce 1931 (obr. 6.24). Generace jsou označeny římskými číslicemi, jednotlivci v dané generaci jsou označeni arabskými číslicemi.
Rýže. 6.24. Legenda při sestavování rodokmenů (podle G. Justa)
Pomocí genealogické metody lze určit dědičnou povahu studovaného znaku a také typ jeho dědičnosti (autosomálně dominantní, autozomálně recesivní, X-vázaný dominantní nebo recesivní, Y-vázaný). Při analýze rodokmenů pro několik charakteristik lze odhalit propojenou povahu jejich dědičnosti, která se používá při sestavování chromozomálních map. Tato metoda umožňuje studovat intenzitu mutačního procesu, posoudit expresivitu a penetraci alely. Je široce používán v lékařském genetickém poradenství k předpovídání potomků. Je však třeba poznamenat, že genealogická analýza se výrazně zkomplikuje, když rodiny mají málo dětí.
Rodokmeny s autozomálně dominantní dědičností. Autozomální typ dědičnosti je obecně charakterizován stejnou pravděpodobností výskytu tohoto znaku u mužů i žen. Je to způsobeno stejnou dvojitou dávkou genů umístěných v autosomech všech zástupců druhu a přijatých od obou rodičů a závislostí vyvíjejícího se znaku na povaze interakce alelických genů.
Když nějaký znak dominuje u potomků rodičovského páru, kde je jeho nositelem alespoň jeden rodič, objevuje se s větší či menší pravděpodobností v závislosti na genetické konstituci rodičů (obr. 6.25).
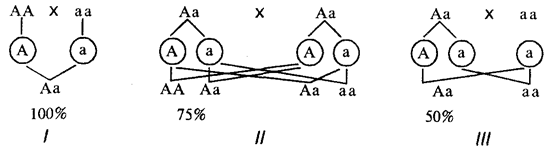
Rýže. 6.25. Pravděpodobnost potomků s dominantním rysem z různých manželských párů (/- III)
Pokud je analyzován znak, který neovlivňuje životaschopnost organismu, pak nositeli dominantního znaku mohou být buď homo- nebo heterozygoti. V případě dominantní dědičnosti některého patologického znaku (onemocnění) homozygoti zpravidla nejsou životaschopní a nositeli tohoto znaku jsou heterozygoti.
U autozomálně dominantní dědičnosti se tedy tato vlastnost může vyskytovat stejně u mužů i žen a lze ji vysledovat, když je v každé vertikální generaci dostatečný počet potomků. Při rozboru rodokmenů je třeba pamatovat na možnost neúplného průniku dominantní alely v důsledku interakce genů nebo faktorů prostředí. Míru penetrance lze vypočítat jako poměr skutečného počtu nositelů znaku k počtu očekávaných nositelů tohoto znaku v dané rodině. Je také nutné pamatovat na to, že některá onemocnění se neprojeví hned od narození dítěte. Mnoho nemocí zděděných podle dominantního typu se vyvíjí až v určitém věku. Huntingtonova chorea se tedy klinicky projevuje do 35.–40. roku života, pozdě se projevuje i polycystická choroba ledvin. Proto se při předpovídání takových nemocí neberou v úvahu bratři a sestry, kteří nedosáhli kritického věku.
První popis rodokmenu s autozomálně dominantním typem dědičnosti anomálie u lidí byl uveden v roce 1905. Sleduje přenos v průběhu řady generací brachydaktylie(krátkoprstý). Na Obr. Obrázek 6.26 ukazuje rodokmen s touto anomálií. Na Obr. Obrázek 6.27 ukazuje rodokmen s retinoblastomem v případě neúplné penetrance.
Rodokmeny s autozomálně recesivní dědičností. Recesivní znaky se fenotypově objevují pouze u homozygotů pro recesivní alely. Tyto znaky se obvykle nacházejí u potomků fenotypově normálních rodičů, kteří jsou nositeli recesivních alel. Pravděpodobnost výskytu recesivního potomstva je v tomto případě 25%. Pokud má jeden z rodičů recesivní rys, pak bude pravděpodobnost jeho projevu u potomků záviset na genotypu druhého rodiče. U recesivních rodičů všichni potomci zdědí odpovídající recesivní rys (obr. 6.28).

Rýže. 6.26. rodokmen ( A) s autozomálně dominantním typem dědičnosti (brachydaktylie - B)
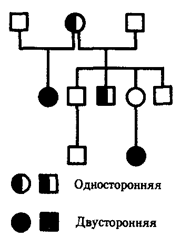
Pro rodokmeny s autozomálně recesivním typem dědičnosti je typické, že se znak neobjevuje v každé generaci. Nejčastěji se recesivní potomci objevují u rodičů s dominantním znakem a pravděpodobnost takového potomka se zvyšuje v blízce příbuzných manželstvích, kde mohou být oba rodiče nositeli stejné recesivní alely přijaté od společného předka. Příkladem autozomálně recesivní dědičnosti je rodokmen rodiny s pseudohypertrofická progresivní myopatie, ve kterých jsou běžná příbuzenská manželství (obr. 6.29). Pozoruhodné je horizontální šíření choroby v poslední generaci.
Rýže. 6.27. Rodokmen s retinoblastomem v případě neúplné penetrace
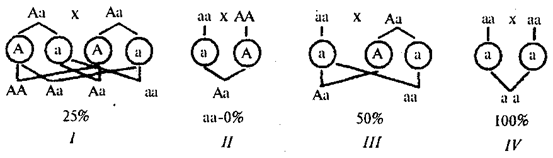
Rýže. 6.28. Pravděpodobnost produkce potomků s recesivním znakem
od různých manželských párů ( I-IV)
Rodokmeny s dominantní X-vázanou dědičností vlastnosti. Geny umístěné na chromozomu X a nemající alely na chromozomu Y jsou přítomny v genotypech mužů a žen v různých dávkách. Žena dostává své dva chromozomy X a odpovídající geny od otce i matky, zatímco muž zdědí svůj jediný chromozom X pouze od své matky. Vývoj odpovídajícího znaku u mužů je dán jedinou alelou přítomnou v jeho genotypu, zatímco u žen je výsledkem interakce dvou alelických genů. V tomto ohledu se znaky zděděné X-vázaným způsobem vyskytují v populaci s různou pravděpodobností u mužů a žen.
U dominantní X-vázané dědičnosti je tento rys častější u žen kvůli větší možnosti, že obdrží odpovídající alelu buď od otce, nebo od matky. Muži mohou tuto vlastnost zdědit pouze po své matce. Ženy s dominantním rysem jej předávají stejnou měrou dcerám a synům, zatímco muži pouze dcerám. Synové nikdy nezdědí dominantní X-spojenou vlastnost od svých otců.
Rýže. 6.29. Rodokmen s autozomálně recesivním typem dědičnosti (pseudohypertrofická progresivní myopatie)
Příkladem tohoto typu dědictví je rodokmen popsaný v roce 1925 s folikulární keratózou - kožní onemocnění doprovázené ztrátou řas, obočí a vlasů na hlavě (obr. 6.30). U hemizygotních mužů je typický těžší průběh onemocnění než u žen, které jsou nejčastěji heterozygotní.
U některých onemocnění je pozorována smrt hemizygotních samců. raná stadia ontogeneze. Pak by v rodokmenech mezi postiženými měly být pouze ženy, v jejichž potomcích je poměr postižených dcer, zdravých dcer a zdravých synů roven 1:1:1. Dominantní mužští hemizygoti, kteří neumírají ve velmi raných stádiích vývoje, se nacházejí při spontánních potratech nebo mezi mrtvě narozenými dětmi. Tyto znaky dědičnosti u lidí jsou charakterizovány pigmentární dermatózou.
Rodokmeny pro recesivní X-vázanou dědičnost vlastností. Charakteristickým znakem rodokmenů s tímto typem dědičnosti je převládající projev znaku u hemizygotních mužů, kteří jej dědí od matek s dominantním fenotypem, které jsou nositelkami recesivní alely. Zpravidla se tento rys dědí muži po generace od dědečka z matčiny strany po vnuka. U žen se projevuje pouze v homozygotním stavu, jehož pravděpodobnost se zvyšuje s blízce příbuznými sňatky.
Nejznámějším příkladem recesivní X-vázané dědičnosti je hemofilie. Dědičnost hemofilie typu A je prezentována v rodokmenu potomků anglické královny Viktorie (obr. 6.31).
Rýže. 6.30. Rodokmen s X-vázanou dominantní dědičností (keratosis follicularis)
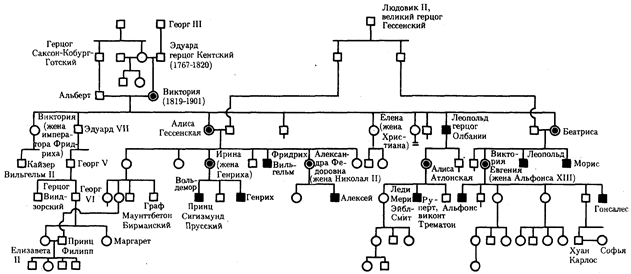
Rýže. 6.31. Rodokmen pro X-vázanou recesivní dědičnost (hemofilie typu A)
Dalším příkladem dědičnosti tohoto typu je barvoslepost - určitá forma poruchy barevného vidění.
Rodokmeny s Y-vázanou dědičností. Přítomnost chromozomu Y pouze u mužů vysvětluje vlastnosti dědičnosti vázané na Y, neboli holandrické, dědičnosti vlastnosti, která se vyskytuje pouze u mužů a je přenášena prostřednictvím mužské linie z generace na generaci z otce na syna.
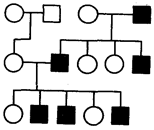
Rýže. 6.32. Rodokmen s Y vázaným (holandrickým) typem dědičnosti
Jedna vlastnost, jejíž dědičnost vázaná na Y u lidí je stále diskutována, je hypertrichóza ušního boltce, nebo přítomnost chlupů na vnějším okraji ucha. Předpokládá se, že kromě tohoto genu obsahuje krátké rameno chromozomu Y geny, které určují mužské pohlaví. V roce 1955 byl u myši popsán transplantační antigen určovaný chromozomem Y zvaný HY. Možná je to jeden z faktorů sexuální diferenciace mužských gonád, jejichž buňky mají receptory, které vážou tento antigen. Antigen asociovaný s receptorem aktivuje vývoj gonády podle mužského typu (viz část 3.6.5.2; 6.1.2). Tento antigen zůstal v procesu evoluce téměř nezměněn a nachází se v těle mnoha živočišných druhů, včetně lidí. Dědičnost schopnosti vývoje gonád podle mužského typu je tedy určena holandrickým genem umístěným na chromozomu Y (obr. 6.32).
Dvojitá metoda
Tato metoda spočívá ve studiu vzorců dědičnosti vlastností u párů jednovaječných a bratrských dvojčat. V roce 1875 Galton původně navrhl posoudit roli dědičnosti a prostředí ve vývoji lidských duševních vlastností. V současné době je tato metoda široce používána při studiu dědičnosti a variability u lidí k určení relativní role dědičnosti a prostředí při utváření různých znaků, normálních i patologických. Umožňuje identifikovat dědičnou povahu znaku, určit penetraci alely a vyhodnotit účinnost některých vnějších faktorů (léky, výcvik, vzdělání) na tělo.
Podstatou metody je porovnat projev znaku u různých skupin dvojčat s přihlédnutím k podobnostem či rozdílům jejich genotypů. Monozygotní dvojčata, vyvíjející se z jednoho oplodněného vajíčka jsou geneticky totožné, protože mají 100% stejné geny. Mezi jednovaječnými dvojčaty je proto vysoké procento souhlasné páry, ve kterém se u obou dvojčat rozvíjí rys. Srovnání jednovaječných dvojčat odchovaných v různé podmínky postembryonální období, umožňuje identifikovat znaky, na jejichž vzniku se významně podílejí faktory prostředí. Podle těchto znaků mezi dvojčaty existuje nesoulad, těch. rozdíly. Naopak zachování podobností mezi dvojčaty i přes rozdíly v podmínkách jejich existence ukazuje na dědičné podmínění znaku.
Srovnání párové shody pro tento znak u geneticky identických jednovaječných a dvojvaječných dvojčat, která mají v průměru asi 50 % společných genů, umožňuje objektivněji posoudit roli genotypu při utváření znaku. Vysoká konkordance u párů jednovaječných dvojčat a výrazně nižší shoda u párů dvojvaječných dvojčat svědčí o důležitosti dědičných rozdílů u těchto párů pro určení znaku. Podobnost míry konkordance u mono- a dizygotních dvojčat ukazuje na nevýznamnou roli genetických rozdílů a určující roli prostředí při utváření znaku nebo rozvoji onemocnění. Výrazně odlišná, ale spíše nízká míra shody u obou skupin dvojčat umožňuje posoudit dědičnou predispozici k vytvoření znaku, který se vyvíjí pod vlivem faktorů prostředí.
Stanovení relativní role dědičnosti a prostředí při rozvoji různých patologických stavů umožňuje lékaři správně posoudit situaci a provést preventivní opatření, pokud existuje dědičná predispozice k onemocnění, nebo provést pomocnou terapii, pokud je dědičná.
Obtíže metody dvojčat jsou spojeny za prvé s relativně nízkou frekvencí porodů dvojčat v populaci (1:86-1:88), což komplikuje výběr dostatečného počtu párů s tímto znakem; za druhé, s identifikací monozygotnosti dvojčat, která má velká důležitost získat spolehlivé závěry.
K identifikaci monozygotnosti dvojčat se používá řada metod. 1. Polysymptomatická metoda pro porovnávání dvojčat podle mnoha morfologických charakteristik (pigmentace očí, vlasů, kůže, tvar vlasů a rysy srsti na hlavě a těle, tvar uší, nosu, rtů, nehtů, těla, vzory prstů ). 2. Metody založené na imunologické identitě dvojčat na základě erytrocytárních antigenů (systémy ABO, MN, Rhesus) a sérových proteinů (γ-globulin). 3. Nejspolehlivějším kritériem pro monozygotnost je transplantační test pomocí cross-twin kožního štěpu.
Přes pracnost metody dvojčat a možnost chyb při určování monozygotnosti dvojčat ji vysoká objektivita závěrů řadí mezi široce používané metody genetického výzkumu u lidí.


















